





From Kerry McNicol 25/03/2013 to Derek Butcher.
Hi Derek, I am pleased that you can take time to help me out.
The plant I have as Nidularium Maggie Hicks is a slow growing (and reproducing) Nid., approximately 30-35 cm across, pale green very tender leaves both on top and under, which mark easily.
The inflorescence consists of greenish bracts turning to pale orange.(this pic isn't as orange as usual)
The petals are greenish with a white lip.
The seed pods are red tipped with a white base(bulb), they make seed very readily.
See Nidularium Maggie Hicks photos below.








Ed - This plant is probably also Nid. amazonicum following W. smithii now being treated as a synonym of amazonicum. |
From Derek Butcher 26/03/13 to Kerry McNicol.
Early in the 1980's Maggie Hick, Josie Tonkin and ourselves grew Wittrockia amazonica from the USA seed bank. When the plants flowered there was similarity but slight differences. All had dull primary bracts and not the bright colour you expect with Nidularium. Anyway since then there has been lots of writings on this species, some 8 pages - see below. So there were problems with identity in the past! BUT we now have the name Nidularium amazonicum. I query the photos on BinA website (1st 4 pictures) but have no proof. I see great similarity with your plant in its two forms with what I think is N. amazonicum. See attached photos and drawings which if you are keen you can use to compare with a flower you have dissected.


Nidularium amazonicum (Baker) Linden & E. Morren ex Lindm., Ofvers. Forh. Kongl. Svenska Vetensk.-Akad. 47 (10): 541.1890. (From Leme in Nidularium – Broms. Atl. Forest 2000
Basionym: Karatas amazonica Baker, Card. Chron. II. 25: 814. 1886.
Synonym: Nidularium amazonicum Linden & E. Morren, Cat. Bromel. 11. Oct. 1873; nom. nud.
Bromelia amazonica Linden ex E. Morren, Cat. Bromel. 11. Oct. 1873; nom. nud.
Aechmea amazonica hort. ex Baker, Card. Chron. II. 25.814. 1886; nom. nud.
Canistrum amazonicum (Baker) Mez, in .Martius, Fl. bras. 3 (3): 249.1891.
Wittrockia amazonica (Baker) L. B. Smith, Arq. Bot. Estado Sao Paulo II. 2. 197. 1952. Syn. nov.
Wittrockia smithii Reitz, Anais Bot. Herb. "Barbosa Rodrigues" 4: 19, pl. 5. 1952. Type: Santa Catarina, Fachinal, 15 Jul. 1951 , R. Reitz 4207 (HBR).
Detail from Leme in Nidularium –Brom. Atl. Forest 167-70. 2000
LEAVES ca. 20, arcuate, densely rosulate, thin in texture, forming a broad funnelform rosette;
SHEATHS elliptic, 12 x 6-7 (-12) cm, densely brown-lepidote;
BLADES narrowly lanceolate, narrowed toward base, 20-50 x 3-6 (-9) cm, canaliculate, bearing a slightly thicker median channel, lepidote toward apex to glabrescent, opaque adaxially, green or very often red-brown or purple abaxially, apex acuminate-caudate, margins very densely and finely spinulose, spines ca. 0.5 mm long, 2-3 mm apart.
SCAPE 5-7 cm long, shorter than the leaf sheaths;
SCAPE BRACTS subfoliaceous.
INFLORESCENCE tripinnate, obconic, capitate-rosulate, ca. 6 cm long, 8-10 (-15) cm in diameter at apex, slightly exceeding the leaf sheaths but not elevated above the rosette;
PRIMARY BRACTS ovate-lanceolate, apex shortly acuminate-caudate, suberect but the apex recurved, reddish-brown to reddish-orange, densely lepidote toward apex, very densely spinulose toward apex, the basal bracts 10-12 x 5-7 cm.
FASCICLES ca. 8, the lower ones 30-35 x 20-25 mm wide, 5-to-8- (13) flowered;
FLORAL BRACTS broadly ovate, apex acute, entire, ca. 25 x 20 mm, membranaceous, sublanate near the apex, about equaling 1/2 to 2/3 sepal length;
FLOWERS 45-56 mm long, nearly sessile;
SEPALS subsymmetrical, oblong-elliptic, apex acute and apiculate, 20-25 x 7-10 mm, connate for 2-7 mm, membranaceous, glabrous;
PETALS 30-35 x 5 mm, greenish white or yellowish with white margins, basal tube 8-10 mm long, bearing 2 fimbriate appendages at apex of tube, plus 2 longitudinal callosities equaling filament length;
FILAMENTS the antepetalous ones almost completely adnate to the petals, the antesepaIous ones adnate to the petal tube then free for 18-20 mm; ANTHERS 5-6 mm long, apex distinctly apiculate; STIGMA subglobose, ca. 1 mm long, margins inconspicuously crenulate;
OVARY ellipsoid-obovate, 10-12 mm long, ca. 6 mm in diameter.
FRUITS white with pale-brown, irregular spots, persistent calyx bright red toward apex.
TYPE: Kew Hortus s. n., fl. cult. 1878. Holotype: K.
ETYMOLOGY
When N amazonicum was published in 1886, it was erroneously thought to have come from the " Amazon Valley" in northern Brazil and this was what inspired the name of this typical Atlantic-forest inhabitant from southern and southeastern Brazil.
DISCUSSION
In the protologue of Karatas amazonica, Baker stated that he had seen the species in flower twice: once at Kew, in 1878, and again in 1886 when he identified a specimen for William Bull. At the first opportunity, Baker did not publish the species since Edouard Morren had already given the taxon a name, "Nidularium amazonicum", and had included the plant under this name (but with no description) in his "Catalogue des Bromeliacees cultivees au Jardin botanique de l'Universite de Liege", in 1873. In the same publication, Morren attributed the name Bromelia amazonica to Jean Jules Linden, who had suggested this epithet. Morren's artist-assistants made a beautiful watercolor of the species which is now a part of the Royal Botanic Gardens collection at Kew.
Only in 1886 did Baker decide to describe the species that Linden introduced to cultivation in the mid 1870s. According to the protologue, this species was already known in horticulture by names such as Aechmea amazonica, as well as those mentioned by Morren (1873). Baker himself (1889) declared that the species had been described on the basis of material that flowered at Kew in 1878, and also based on Bull's material from June 1886 (in that order). However, only the first material is compatible with the exsiccata in Kew and this must necessarily be considered the holotype of N.amazonicum. It is composed of one leaf and several flowers. There are no exsiccata that correspond to the second material mentioned by Baker The drawing that is attached to the holotype was produced by Morren based on a specimen deposited at the herbarium of the University of Liege, dated October 1874. The fruiting structure of this specimen has practically no primary bracts. Baker only saw this drawing at Kew in late 1888 (Baker, 1889) and it is not part of the protologue which is dated 1886.
In 1890, Lindman clearly referred to Morren's (1873) catalogue and revalidated the name Nidularium amazonicum as a new combination for Baker's basionym, attributing it to Linden and Morren. Mez (1891) transferred the species to the genus Canistrum and later Smith (1952) included it in Wittrockia. When it was transferrd to Wittrockia, Smith (1952) confirmed the type to be the material that flowered in the Royal Botanic Gardens, Kew, in 1878. However, the probable clonotypes indicated by Smith & Downs (1979) and expanded by Leme & Brito (1993) should be regarded with extreme caution, and were therefore excluded here. Although the stock of specimens of this taxon was probably very small in Europe in the late 19th century, the exsiccata of these alleged clonotypes do not have enough information to positively associate them with the data in the protologue and the holotype material (excluding the sketch by Morren). Furthermore, Baker (1889) stated that Morren had written on the painting of the species that the material (probably that which produced the exsiccata at Liege) had been sent from "Rio" (i.e., Rio de Janeiro) by Binot, which also does not agree with the information contained in the protologue.
More recently, Leme & Brito (1993) considered Wittrockia smithii to be a synonym of W. Amazonica because there were no consistent characteristics to differentiate Reitz's species from what was then known as W. Amazonica. In fact, it was merely a case of inconsistent terminology used in the morphological description of the respective type specimens, which, in themselves, were not substantially different.
It is interesting to note that the exsiccata that flowered in April 1873 and January 1874 in Liege were identified at the time, probably by Morren, as N. innocentii, thus representing the onset of the conceptual chaos that has reigned until today.
Nidularium amazonicum and N. innocentii, both with wine-colored leaves abaxially, were often mistaken for one another. Foster (1960) tried to solve the problem by indicating the historical motives that led to confusion of the two taxa. However, the state of chaos endured due to the absence of a precise conceptualization of N. amazonicum, which most certainly is why Reitz published the synonym W. Smithii.
Although N. innocentii and N. amazonicum are both part of the same "white complex" of species, they belong to different subcomplexes. The petals of N. innocentii form a well-developed basal tube with totally white lobes and no appendages. In contrast, N. amazonicum has petals that are shortly connate at base, greenish white or yellowish with white margins and also with well-developed fimbriate appendages.
Nidularium amazonicum shows a clear morphological affinity to N. krisgreeniae. It is distinguished by its acuminate-caudate leaf blades without transverse lines, inflorescence sunken in the leaf rosette, primary bracts suberect-recurved, and sessile flowers. In the wild, N. amazonicum allows a color-variation pattern. Some populations have totally green leaves while others are purplish wine especially on the abaxial leaf surface, a trait more common in Santa Catarina. The primary bracts vary from reddish brown to red or reddish orange. When exposed to more sunlight, the inflorescence of N. amazonicum is deeply sunken in the leaf rosette, while in shadier spots in the forest it is more protruding, but it never rises above the leaf rosette as in N. krisgreeniae. The corollas of this taxon also vary in color pattern from whitish green to yellowish (yellowish-green) with white margins.
DISTRIBUTION & HABITAT
Nidularium amazonicum is concentrated geographically in the states of southern Brazil (Parana, Santa Catarina and, more diffusely, Rio Grande do Sul ). There is only one collection recorded from Sao Paulo. The species lives in the Atlantic slope forest from sea level to ca. 900 m altitude. It grows mainly on the ground and on rocks, but also occurs occasionally as an epiphyte in the lower layer of the forest. It flowers from February to April and is represented in cultivation by at least five distinct clones.
In Parana and Santa Catarina, N. amazonicum is sympatric with N. innocent and N. procerum, while in Rio Grande do Sul is grows together with N. jonesianum.
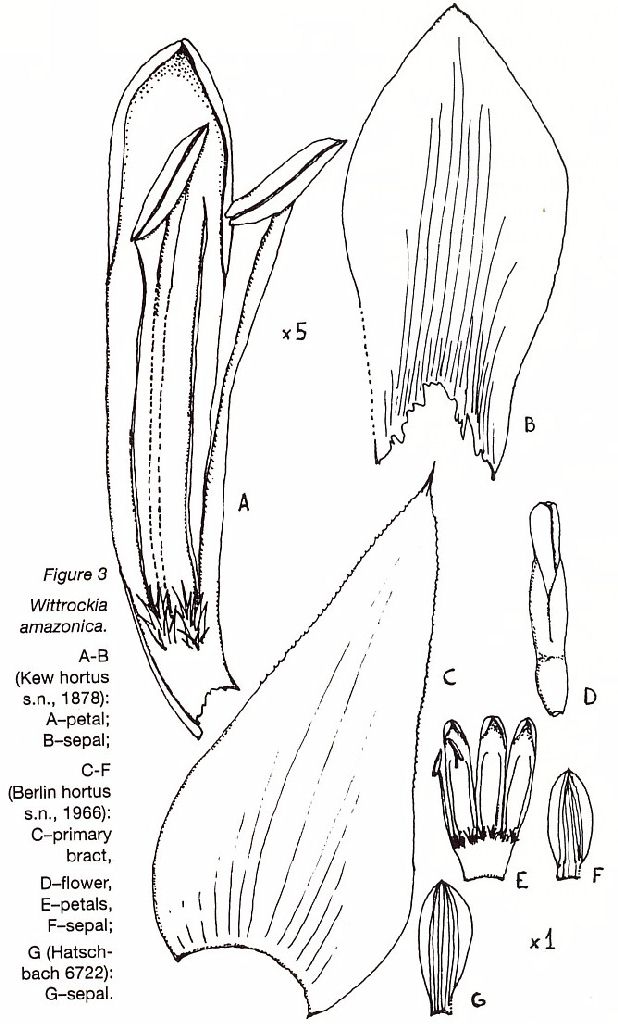
Wittrockia amazonica; A Long-Standing Mystery Unveiled by Elton M.C. Leme in J. Brom. Soc. 43(1):3-6, 32. 1993
Introduced into cultivation by Linden in 1870, Wittrockia amazonica played an outstanding role in horticulture toward the end of the last century. A flowering specimen was seen for the first time by J.G. Baker in 1878 although he did not describe it officially until 1886 since he had expected Edouard Morren to do that. At the time, it was believed that the species originated in the "Valley of the Amazon," which gave rise to its provisional name of Nidularium amazonicum, originally attributed by Linden and Morren in 1873.
The species was transferred to the genus Wittrockia by Lyman B. Smith in 1952 because of the appendages at the bases of its partially connate petals. The curious point is that all known specimens in herbaria (a total of four) are reputed to be clonotypes, meaning that they came from the same source of cultivation of the type specimen deposited in the Royal Botanic Gardens, Kew. We have not heard, however, of any other descendants of the original clone that have survived to the present day in cultivation.
The fact that we have never located Wittrockia amazonica in either the wild state or in Brazilian herbaria has always intrigued us, particularly when we considered that the genus is endemic to the region of Brazil where we have concentrated our surveys and research projects. But finally, having examined the specimens from Kew and the Berlin-Dahlem Botanical Museum, as well as photocopies of the specimens from Liege University in Belgium and the Smithsonian Institution in Washington, D.C., we believe that we have the solution to this mystery.
In accordance with the key drawn up by Smith & Downs, Wittrockia amazonica is shown to have affinities with W. smithii Reitz. The basic differences are the free sepals as well as secondary characteristics such as the shape of the sepals and the degree of concrescence of the petals of W. amazonica. In addition, these characteristics do not appear either expressly or implicitly in the original description of that species.
That being the case, we move on to an examination of its closest relative, Wittrockia smithii with over twenty dried specimens in Brazilian herbaria including the Herbarium Bradeanum, Instituto de Botanica de Sao Paulo, Museu Botanica Municipal de Curitiba, and Herbario Barbosa Rodrigues. Those specimens came from a wide range of areas including the states of Sao Paulo, Parana, Santa Catarina, and Rio Grande do Sul. In addition, we collected specimens in the states where the species is most common, Parana and Santa Catarina, and made various observations largely on the variability of this species. As the result of these studies it is now possible to outline the average characteristics of W. smithii:
Sepals no more than acute and connate reveal connation at their base of about 2-3 mm.
Petals uniformly connate at base for about 10 mm, varying in color including greenish white, green with white margins, yellowish with whitish margins.
Leaf color varying from pure green to reddish brown or purplish mainly on the underside.
As the dimensions and the morphological standard of the leaves and inflorescences of both species seem identical to us, W. smithii would be differentiated from W. amazonica only if, in accordance with the concept of the Smith & Downs monograph, the sepals of the latter were in fact free, strongly asymmetrical, and acuminate. Since, however, we were able to study the original material of W. amazonica, despite its fragmentary state, as well as other original material from a probable clonotype in perfect condition, we noted that its sepals are basally connate for 2-3 mm, slightly asymmetrical and no more than acute at apex. For these reasons, we do not confirm the small, but important, alterations made over the course of time in the original concept of W. amazonica, and conclude that there is no significant morphological element that differentiates it from W. smithii. The latter should, therefore, be reduced to synonomy as follows:
Wittrockia amazonica (Baker) L.B. Smith, Arq. Bot. S. Paulo II. 2:197.1952.
Nidularium amazonicum Linden & Andres, Cat. Linden 11. 1873; nomen.
Karatas amazonica Baker, Gard. Chron. IL25:814. 1886.
Bromelia amazonica hortus ex Andre, Revue Hort. 58:503. 1886; nomen.
Aechmea amazonica hortus ex Andre, Revue Hort. 58:503. 1886; nomen.
Nidularium amazonicum (Baker) Lindman & E. Morren, Oefvers Vet. Akad. Forhandl. 47:541. 1890.
Canistrum amazonicum (Baker) Mez, Mart. Fl. Bras. 3(3):249.1891.
Wittrockia smithii Reitz, Anais Bot. Herb. Barbosa Rodrigues 4:19, p1.5. 1952. Type. Fachinal, Santa Catarina, Brazil, R. Reitz 4207 (HBR), 15 July. 1951.
Plant epiphytic or saxicolous, stemless, propagating by short basal rhizomes.
Leaves 15-20 in a dense rosette, arcuate, submembranaceous or papyraceous;
sheaths elliptic, ca. 12 cm long, 6-7 (-12) cm wide, densely brown-lepidote on both sides, abaxially often tinged with red-brown or purple; blades narrowed toward base, narrowly lanceolate, acuminate-caudate, canaliculate with a more rigid median channel, 20-50 cm long, 3-6 (-9) cm wide, very densely and finely serrulate;
spines ca. 0.5 mm long, green or very often tinged abaxially with red-brown or purple, lepidote to glabrescent toward apex;
Scape 5-7 (-14) cm long; scape bracts subfoliaceous;
Inflorescence densely bipinnate to inconspicuously tripinnate, subcorymbose, subcapitiform-obconic, sunk in the rosette, ca. 6 cm long, ca. 10 (-15) cm in diameter at apex;
primary bracts oval-lanceolate, apex short acuminate-caudate, ca. 10 (-12) cm long, ca. 5 (-7) cm wide at base, reddish brown to bright red, densely lepidote toward apex, very densely serrulate, suberect but the apex recurving;
fascicles flabellate, complanate, with 3-8 (-13) flowers, 30-35 mm long (without the petals);
floral bracts ovate, to nearly equaling the center of the sepals, acute, entire, lepidote, carinate, membranaceous;
flowers 40-50 (-56) mm long, near sessil;
sepals oblong-elliptic, apex acute and apiculate, subsymmetric, ca. 20 (-25) mm long, 7-10 mm wide, connate at base for 2-3 (-7) mm, glabrous;
petals linear, 30-35 mm long, apex obtuse-cucullate, connate at base for 10 mm, bearing 2 fimbriate appendages ca. 10 mm above the base and 2 longitudinal calluses equaling the length of the filaments, blades erect, greenish white, green or yellowish with whitish margins; filaments: the epipetalous ones highly adnate to the petals, the episepalous adnate for ca. 10 mm;
ovary 10-12 mm long, ca. 6 mm in diameter, ellipsoid-obovate, slightly constricted at apex;
placentae subcentral; epigynous tube ca. 1 mm long;
ovules many, obtuse.
Basic material studied:
Holotype: Kew Hortus s.n., flowered in 1878 (K).
Probable clonotypes: Kew Hortus s.n., Sept. 1885 (K); Liege Hortus s.n., Oct. 1874 (LG); DeLeon s.n., 1 Nov. 1961 (US); Berlin Hortus s.n., 10 Mar. 1966 (B).
Diverse material: Brazil, Santa Catarina State, Antonio Carlos (old Biguacu), Fachinal, R. Reitz 4207,15 July 1951 (HBR).
The reason for the "disappearance" of Wittrockia amazonica from the wild now becomes quite understandable. With the stated characteristics, principally those of its sepals, misinterpreted it could never have been recognized except as W. smithii, which from the 1950s onwards acted as a disguise for its identity and forms the key piece in this puzzle.
NOMENCLATURE CLARIFICATION No. 11 by Mulford B. Foster in BSBull. 10(4): 56,58. 1960
Nidularium amazonicum vs. Wittrockia amazonica
Feeling that I have been partly responsible for the confusion, at least in the United States, which has been caused by the mislabeling of a popular Nidularium which has been sold in this country for the past twenty-five years, it is high time that something was done to correct the situation.
For more than twenty years I have had a plant in my collection, labeled. It originally came to the U. S. from Europe where it had been so labeled for many years. This Nidularium, which has metallic magenta-green leaves, glossy beneath, with a rather pronounced sunken midrib, has a central inflorescence consisting of rusty red bracts, each containing a cluster of white flowers. This inflorescence is in the center of the rosette of leaves and its proper name is Nidularium Innocentii Lem var. Innocentii (named for M. St. Innocent and NOT Wittrockia amazonica as many persons have recently, wrongly, relabeled it.
When Dr. Lyman B. Smith published, in 1955, THE BROMELIACEAE of BRAZIL, many of our bromeliad enthusiasts saw, for the first time, the complete list of known Wittrockia species as shown on pages 185-186, Under Wittrockia amazonica (Baker) L. B. Smith, three synonyms are listed. Baker called this plant Karatas amazonica. Linden and Morren called it Nidularium amazonicum while Mez named it Canistrum amazonicum.
This plant was cultivated and flowered in the Kew Gardens, London, in 1878 and was listed in their Hand-List as late as 1915. At that time practically all of the species then listed as Karatas are now known as species of either, Nidularium, Neoregelia or Wittrockia. Morren made a drawing of this "Karatas amazonicum” and stated on the drawing that it was received from Mr. Wm. Bull in June 1886 and that it came from Binot Rio de Janeiro. Baker states that it was introduced by Linden about 1870.
How long this "Wittrockia amazonica" lived under its different generic names, we do not know, nor do we know when the label of “Nidularium amazonicum” was mistakenly shifted (in some distant greenhouse) over on to the plant of what has been legally, botanically determined Nidularium innocentii. But we do know that, now, at the present time, a number of Bromeliad Society members have placed another wrong label of "Wittrockia amazonica” on the plant which does not even belong to Wittrockia, neither is it "Nidularium amazonicum", but is in truth, Nidularium innocentii var. innocentii.
The photo of the true Nidularium innocent var. innocentii, above, was taken at Alto de Serra in Sao Paulo, Brazil when we were guests on that inspiring reservation in 1939.
It is to be hoped that every one of the Society's members will properly label this beautiful plant with its proper name, NIDULARIUM INNOCENTII var. INNOCENTII and NOT "Wittrockia amazonica", nor "Nidularium amazonicum."
While we are on the subject of Nidularium Innocentii, it will be helpful to list the five varieties of this species after the clarification in Lyman Smith's BROMELIACEAE of BRAZIL on page 167. (The seeming double-talk of the first species name, to indicate that it is the first one discovered; all the others have been found since, and are variations of the first one.)
Plants with dark, magenta-green leaves: var. innocentii
Plants with all green leaf blades: var. wittmackianu
Plants with green leaf blades marked with longitudinal white lines: var. striatum
Plants with green leaf blades marked with numerous white lines: var. lineatum
Plants with green blades having a single large median white stripe: var. paxianum
(Ed. Note. Paxianum “large median white stripe” has now been dropped from the definition. See discussion on this web site.)
Detail from S&D
4. Wittrockia amazonica (Baker) L. B. Smith, Arq. Bot. S. Paulo II. 2: 197. 1952. Nidularium amazonicum Linden & Andre, Cat. Linden 11. 1873; nomen (cf. under Karatas amazonica Baker).
Karatas amazonica Baker, Gard. Chron. 11. 25: 814. 1886.
Bromelia amazonica hortus ex Andre, Revue Hort. 58: 503. 1886; nomen.
Aechmea amazonica hortus ex Andre, Revue Hort. 58: 503: 1886; nomen.
Nidularium amazonicum (Baker) Lindman & E. Morren, Oefvers. Vet. Akad. Forhandl. 47: 541. 1890.
Canistrum amazonicum (Baker) Mez, Mart. Fl. Bras. 3(3): 249. 1891.
Plant stemless but propagating by rhizomes.
Leaves 15-20 in a dense rosette, 5 dm long, submembranaceous or papyraceous;
sheaths broadly elliptic, lepidote especially toward base;
blades ligulate, acuminate, narrowed toward base, 45 mm wide, densely and finely serrulate, brownish green above with a pale median channel, tinged with red-brown beneath, the outer ones sparsely pale-lepidote or soon glabrous above.
Scape very short;
scape-bracts subtriangular-ovate, exceeding the inflorescence, the same color as the leaves.
Inflorescence densely bipinnate, capitiform;
primary bracts like the scape-bracts;
spikes few-flowered, subfasciculate.
Floral bracts oblong, exceeding the ovaries, colorless, membranaceous, subglabrous;
flowers ca 45 mm long.
Sepals free, strongly asymmetric, acuminate, mucronate, 2 cm long, glabrous;
petals obtuse, cucullate, short-connate, bearing 2 fimbriate scales above the base, blades erect, greenish white;
second series of stamens highly adnate to the petals;
pollen biporate;
ovary subglobose, constricted at apex;
placentae central, the epigynous tube evident;
ovules obtuse.
Type. Kew Hortus s n (holotype, K; photo GH), Brazil.
Distribution. Known from the type collection only. Probable clonotypes: Liege Hortus s n (LG); 1 Nov 1961, De Leon s n (US); 10 Mar 1966, Berlin Hortus s n (B)
5. Wittrockia smithii Reitz, Anais Herb. Barbosa Rodrigues 4: 19, pl. 5. 1952.
Fig 579 A-G.
Leaves about 16 in a spreading rosette, 6 dm long;
sheaths distinct, suborbicular, entire, broadly membranaceous-margined, lepidote on both sides especially toward base;
blades ligulate, acute, filiform -apiculate, contracted at base, 3-6 cm wide, regularly and subdensely serrulate with antrorse curved spines nearly 1 mm long, channeled, undulate along margin, lepidote on both sides, green above, often purple-brown beneath.
Scape almost none.
Inflorescence sunk in the center of the rosette, subcorymbose-compound, depressed-pyramidal, many-flowered;
primary bracts foliaceous, dark red, exceeding the inflorescence;
branches 8-flowered.
Floral bracts obovate, mucronulate, 30 mm long, slightly exceeding the centers of the sepals, carinate, membranaceous, white, entire, subglabrous;
flowers sessile, 56 mm long.
Sepals subsymmetrical, oblong-elliptic, rounded and apiculate, 20 mm long, connate for 7 mm, convolute, greenish white toward base, reddish toward apex;
petals erect, obtuse, cucullate, free at base, connate a little above and then free again, bearing 2 fimbriate scales 10 mm above the base, white with greenish apex; stamens included, filaments of the second series adnate to the petals for most of their length;
anthers 6 mm long;
pollen biporate;
ovary angled, 12 mm long, 6 mm in diameter, white, glabrous;
placentae slightly above center of cell;
ovules obtuse.
Type. Reitz 4207 (holotype, HBR), Fachinal, Biguacu, Santa Catarina, Brazil, 15 Jul 1951.
Distribution. Epiphytic, saxicolous, and terrestrial, 250-900 m alt, Parana and Santa Catarina, Brazil.
BRAZIL. Parana: Curitiba to Paranagua, 15 Sep 1953, Reitz 5760 (HBR, US); Rio Cotia, Antonina, 24 Mar 1966, Hatschbach 14143 (MBM, LS); Serra Capivari Grande, Campo Grande do Sul. 20 Mar 1967, Hatschbach 16207 (MBM, US). Santa Catarina: Ribeirao Grande, Taio, 14 Aug 1951, Reitz 4150 (HBR); Vargem do Cedro, Imarui, 2 Jan 1952, Reitz 4532 (HBR); Anitapolis, 27 Dec 1951, Reitz 4536 (HBR); Morro Spitzkopf, Blumenau, 20 Mar 1952, Reitz 4658 (HBR); L. B. Smith & Reitz 5282 (US); 6292 (R, US); Morro do Bau. Ilhota, 23 Dec 1967, Lourteig 2375 (P, US).