Gardeners
A three part article presented to the
By Don Beard
Far North Coast Bromeliad Study Group N.S.W.
Australia.
A three part article presented to the
By Don Beard
Far North Coast Bromeliad Study Group N.S.W.
Australia.
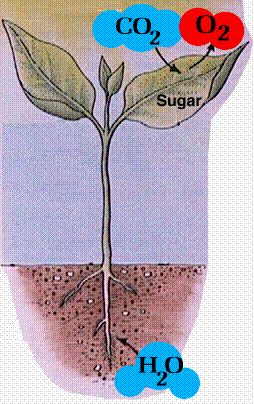
Photosynthesis means the putting together with light or the making or manufacturing with light. It converts light energy into chemical energy and stores it as sugar. It occurs in green plants and requires the green pigment chlorophyll. It is the source of energy for nearly all life, the exception being bacteria and archaea living in extremely hostile environments (chemoautotrophs). This very important but simplified process can be written chemically as:
i.e. green plants in the presence of light combine carbon dioxide and water to make sugar and oxygen. In other words green plants make their own food. This most important equation is the ultimate source of all carbon in the atmosphere.
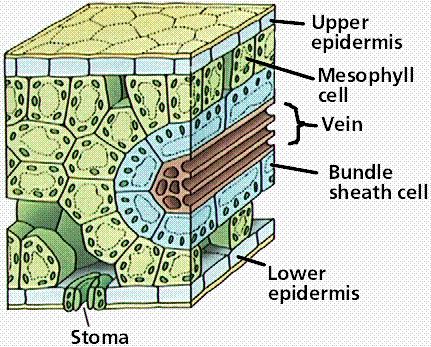
For this process green plants capture approximately100 terawatts of energy in any one year. This is approximately six times the entire human power usage in one year. But where in the plant does this all take place? The answer is in the leaves so we better look more closely at a typical leaf cross-section (see below).
 .
. 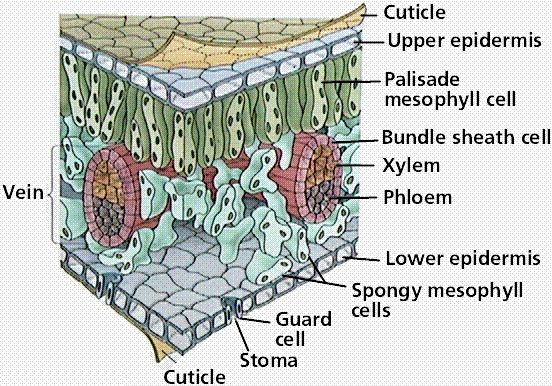
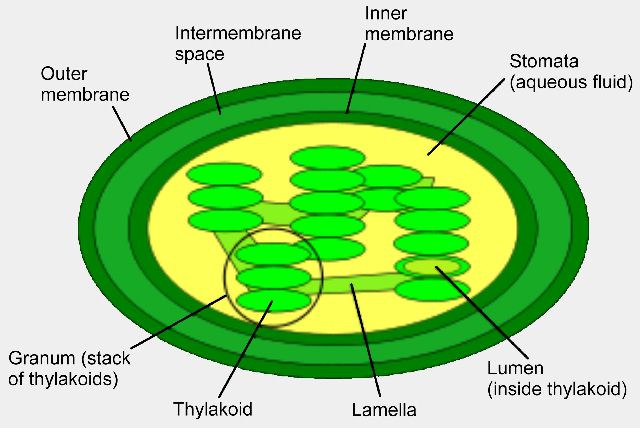
The green colouration occurs in bodies called chloroplasts which in turn occur in the mesophyll of the leaf, not in the surface of the leaf. These chloroplasts contain bodies (thylakoids) which are stacked like pancakes. It is in the margins of these pancakes that the chlorophyll and other pigments occur. It is here that the energy from the sun is absorbed.
Simplified, photosynthesis can be divided into two stages:
Stage 1: or the Light dependant stage is where energy is absorbed from sunlight by the chlorophyll in the Thylakoid membranes. Only the red and blue ends of the spectrum is absorbed and stored. Green light is not used and is reflected, hence the plants green appearance. At this stage oxygen is released.
 .
. 
Stage 2: or the Dark Stage does not require sunlight. This is where the stored energy from the Light Stage is used to convert carbon dioxide and water into an organic compound which has three carbon atoms, a C3 molecule. This reaction occurs in the aqueous fluid in the chloroplast. After six of these cycles glucose is produced. This is the food and energy for the plant. This is also called the Calvin cycle (after Melvin Calvin) and these sorts of plants are referred to as C3 plants and this the C3 pathway. This is a distinctly different pathway to that followed by C4 and CAM plants.
Other Interesting Snippets of Photosynthesis.
The basic process for photosynthesis was understood early in the 18th century. However some stages of photosynthesis are still not fully understood.
If the carbon dioxide level is too low, oxygen will replace it and carbon will be lost from the cycle, sugars drained, toxins produced, and photosynthesis inhibited. The plant will die.
Generally plants sleep at night when their stoma are closed. No carbon dioxide in, no oxygen out.
A typical plant mesophyll cell contains between 10-100 chloroplasts. An area of one square mm of a leaf contains up to 800,000 chloroplasts.
One hectare of corn (which is actually a C4 plant) will produce enough oxygen in one day in mid summer for 325 people.
This presentation has been gleaned from the following internet pages. For those seeking a more detailed and less simplified explanation / introduction to photosynthesis, these same references can be used.
www.biology.clc.uc.edu/courses/bio104/photosyn.htm - (Ed. this link is no longer valid)
(Ed. but this is another good source of information - www.kremp.com/kids-photosynthesis-guide)
www.en.wikipedia.org/wiki/Photosynthesis
www.emc.maricopa.edu/faculty/farabee/biobk/biobookps.html
Photosynthesis 2 by Don Beard This is the second talk by Don in a series of three. The initial talk (see Beard, D. 2012 Photosynthesis 1. FNCBSG NSW Newsletter, April: pp 6-7) introduced the Study Group to basic photosynthesis and the C3 pathway.
We will recall that the equation for photosynthesis, simply expressed, is as above. However one additional item needs to be introduced with respect to C3 plants, and that is the enzyme/catalyst RuBisCo. This is probably the most abundant protein on earth and is used to fix or trap carbon dioxide (CO2) in the process of photosynthesis. In a C3 plant where the first product of photosynthesis is a molecule with three Carbon (C) atoms, RuBisCo acts alone.
Slightly up the evolutionary ladder are plants where the first product of the photosynthetic process is a molecule with four C atoms and where RuBisCo does not act alone. These are C4 plants and were developed along a number of parallel evolutionary lines in order to tolerate aridity, high temperatures and low CO2. These C4 plants developed by some five to 10 million years ago, late in the Miocene. This was also during a maximum glacial period. These plants were all phylogenetically derived from C3 plants. Examples of C4 plants include grasses, maize, corn, sugarcane, sorghum and lots of weeds.
Leaf Anatomy
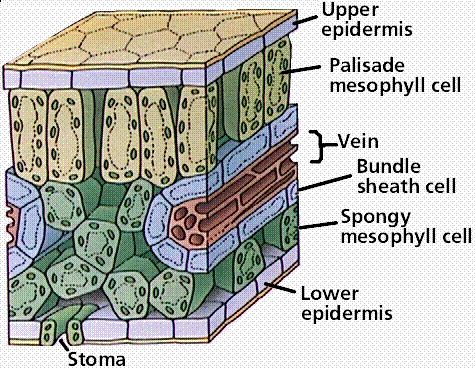
There are anatomical differences between the leaves of C3 plants and C4 plants. Note the differences between the following two drawings (Fig. 6,7).
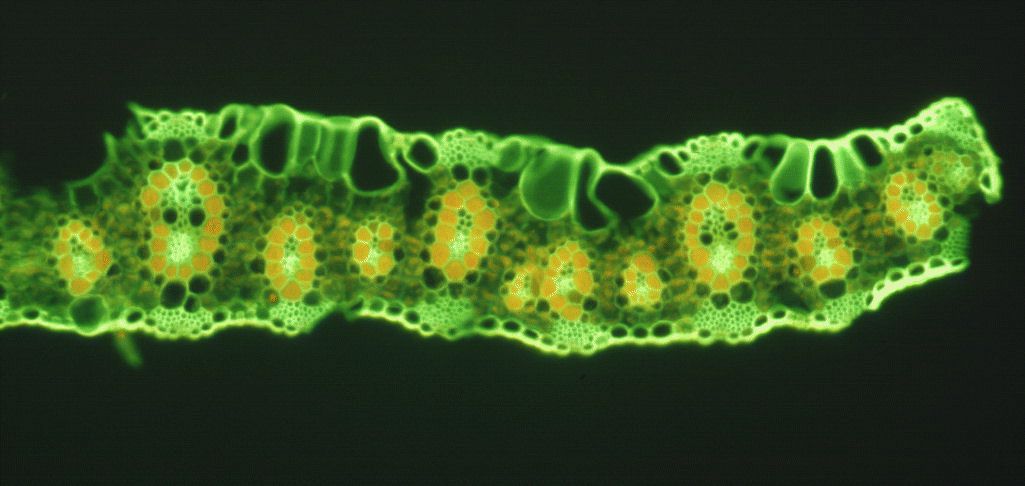
With regard to the vein or vascular bundle, the C4 leaf has a vein that is surrounded by thick walled parenchyma cells which are more tightly packed than for the C3 leaf. These are the bundle sheath cells (BSC) and in a C4 plant it is where the photosynthesis takes place (see photo p13). The much less tightly packed arrangement for the C3 leaf is what eventually allows CO2 to escape back into the atmosphere i.e. the process of photorespiration. This process is negligible to absent in the C4 leaf.
C4 plants generally exhibit parallel venation and have more veins per unit area.
 .
. 
The C4 Mechanism
Whereas the RuBisCo in the C3 plant fixes the CO2 (rather poorly) and prepares for the photosynthesis process in all the chloroplasts in all the mesophyll cells, the C4 plant has a more efficient way of fixing the CO2.
It has a much more efficient enzyme called PEP which compared to RuBisCo has a much greater affinity with CO2. When the stomata open in the morning, the PEP combines with the incoming CO2 and forms oxaloacetic acid and then malic acid. Both these compounds have four carbon atoms in their makeup ... hence the C4 pathway or C4 plant. The malate then travels to the bundle sheath cells (BSC) where it is converted back to CO2 and PEP. The CO2 is then fixed by the RuBisCo in the bundle sheath cells, and photosynthesis occurs with its resultant sugar via the C3 pathway and the Calvin cycle.
The direct C4 pathway...
CO2 + PEP --> oxaloacetic acid --> malic acid --> to bundle sheath cells
Then in bundle sheath cells, in the presence of RuBisCo, using the Calvin cycle...
CO2 + PEP --> photosynthesis --> Sugar
The combined efficiency of PEP in fixing CO2, together with the tightly packed double ring of bundle sheath cells and mesophyll cells (called Kranz anatomy..... meaning ‘wreath’), makes for an easy method of concentrating CO2 without allowing it to escape. An efficient sugar making process. A marked contrast to the C3 plant. An additional feature of the C4 plant is its ability to close it’s stomata in the heat of the day. This of course prevents loss of water. So with low transpiration, negligible photorespiration, and efficient sugar making we have evolved our drought, heat and low CO2 tolerant plant. Note that photorespiration which is in general caused by the uptake of O2 (oxygen) instead of CO2 by the RuBisCo enzyme, undoes the good work of photosynthesis in the C3 plant. From the increased light use efficiency of the C4 plant we improve the quantum yield or in other words growth of the plant. As a consequence of this, many C4 plants are grown commercially and are recognized as some of the world’s major crops.
C4 Plants: Occurrence and History
As stated before C4 plants include many grasses and sedges, many weeds including crabgrass and nutgrass. Also corn, sorghum, millet, sugarcane, and salt
bush.
C4 plants make up 4% of the worlds plant biomass, 15% of all plant species, and 20% of plant commercial production. C4 plants are common as monocots (one seed leaf), 50%, and uncommon as dicots (two seed leaves), 0.6%. Some plants are intermediate between C3 and C4 pathways,i.e. C3 plants exhibiting C4 traits. Some young plants can switch from C3 to C4, some C3 plants have C4 characteristics in their roots, stems, and petioles. Obviously one is not draw- ing too long a bow to think only minor adjustment was needed for a C3 plant to evolve into a C4 plant.
Recent earth history describes a decreasing CO2 level. During the Cretaceous (some 130 million years ago) CO2 was at a level four to five times that of today. This level seriously decreased in the late Oligocene (25-30 million years ago) and continued decreasing to the end of the Miocene (5-10 million years ago) to about 400ppm (parts per million), a little more than today’s level. Under these conditions C4 photosynthesis has developed a number of times in a number of plant lines in the 25-30 million years since the late Oligocene, getting to today’s numbers by the end of the Miocene.
Assuming that low CO2 is a pre-condition for the development of C4 plants, paramaters such as increasing aridity, high light habitats, increasing temperature and seasonality, fire, and the distribution of grazing animals, are all thought to play an important part in this evolutionary trend.
At temperatures 22C - 30C, Quantum yields for C3 and C4 plants are the same
Temperatures above 30C, quantum yields are greater in C4 plants
Temperatures below 22C, quantum yields are greater in C3 plants.
| - Can shut stomata in heat of day | - Can’t |
| - First product has 4 C atoms | - First product has 3 C atoms |
| - PEP and RuBisCo | - RuBisCo only |
| - Tight gas barrier about BSC | - No barrier |
| - BSC have chloroplasts | - BSC have no chloroplasts |
| - Venation... parallel and closer. | - Venation… anything |
| - PEP loves CO2 and wont take up O2 | - RuBisCo can’t tell difference between CO2 and O2 |
| - CO2 absorbed and used fast | - CO 2 absorbed and used slowly |
| - Photosynthesis restricted to BSC chloroplasts | - Photosynthesis operates in all mesophyll chloroplasts |
| - No photorespiration | - Up to 30% photorespiration |
| - Can utilize low CO2 | - Needs high CO2 |
| - High rates of photosynthesis and growth particularly in tropics. Drought tolerant. | - Lower rates of photosynthesis .Can’t handle arid situations and high temperatures. |
| - Dominate open hot arid environments. | - Low water usage efficiency. |
On a final note, rice is a C3 plant. Science has for some years been striving to develop it into a C4 plant. Imagine what that might do for rice production and the world’s food problems.
References: As with Photosynthesis 1, this presentation was gleaned from the following scientific articles and internet pages:
Sage et al, 2011, The C4 plant lineages of planet Earth. J. Exp. Bot.
Sage 2004, The evolution of C4 photosynthesis.
New Phytol. 161: 341-370
www.en.wikipedia.org/wiki/C4_carbon_fixation
www.marietta.edu/~spilatrs/biol103/photolab/c4photo.html
www.creation.com/c4-photosynthesisevolution-or-design
www.ehleringer.net/Jim/Publications/271.pdf
www.plantsinaction.science.uq.edu.au/edition1/?q=content/2-1-5-c4-photosynthesis
www.life.illinois.edu/govindjee/Part1/Part1_Hatch.pdf
www.oregonstate.edu/instruct/css/330/two/
www.en.wikipedia.org/wiki/Photosynthesis
www.emc.maricopa.edu/faculty/farabee/biobk/biobookps.html

Photosynthesis 3 by Don Beard
This is the final talk in a series of three on photosynthesis. An alternative title may well have been “ Photosynthesis for Bromeliad Gardeners”. Previous articles can be seen in FNCBSG(NSW) Newsletters Apr. 2012,pp 6-7; July 2012, pp10-14. In this article the CAM photosynthetic pathway and CAM plants are
discussed. CAM is an acronym for Crassulacean Acid Metabolism, meaning the type of acid metabolism found in the Crassulaceae, a family of succulent plants.
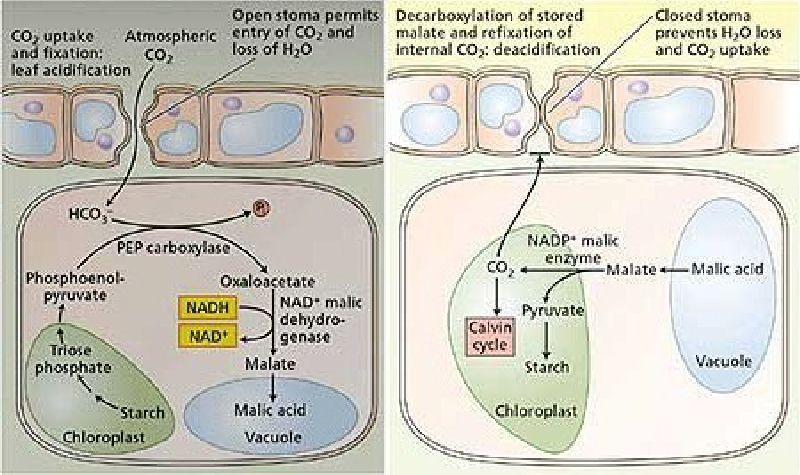
It was developed as an adaptation to arid conditions. Briefly the CAM pathway involves the plant shutting stomata during the day to reduce water loss, opening them at night to collect CO2, and storing the CO2 as the 4C molecule malic acid. Then the next day with the stomata shut, CO2 is reproduced and used for photosynthesis. The malic acid gives the leaf of the CAM plant a bitter/acid taste during the night which disappears during the day.
The term CAM is generally attributed to Thomas and Ransom in 1940, but 2000 years ago the romans noticed the distinctive acid taste that CAM leaves have at
night. However it wasn’t until the early 1930’s that the process was suspected, and then verified during the 1940’s. The process was almost completely understood by1980. Examples of CAM plants include bromeliads, orchids, cacti and Jade plants. Most are epiphytes or succulents.
Mechanism
CAM probably developed as a two part (day/night) 24 hour cycle as an adaptation to increased water efficiency. At night during lower temperatures the stomata open and atmospheric CO2 enters and is fixed in the spongy mesophyll cells by an enzyme reaction (PEPC) forming HCO
Night CO2 --> HCO
At dawn the stomata close, the malic acid moves from the vacuoles, is converted to malate and decarboxylated in the chloroplasts into CO2 and PEP. The CO2 concentrates around the enzyme RuBisCo and photosynthesis via the Calvin cycle results.
Day Malic acid --> Malate decarboxylated --> PEP + CO2 (for Calvin cycle)
In the late afternoon the stomata open and this day/night cycle repeats.
The water efficiency of this process is demonstrated by the fact that C3 plants lose 97% of their water by transpiration whereas CAM plants loose little to none. All this is achieved by just shutting the stomata during the day.
CAM Types
Obligate (Constitutive). Night uptake of CO2 occurs at all times i.e. only the CAM photosynthetic pathway is used by the plant.
Inducible (Facultative). These plants only use CAM when stressed, and can switch from C3/C4 to CAM, depending on the environment.
CAM Cycling. With these plants the stomata don’t open at night. The plants have to recycle the CO2 produced by respiration. These are usually well watered lants that keep their stomata open during the day. Benefits of this type of CAM are not at all obvious. This may be a precursor to CAM Idling.
CAM Idling. This photosynthetic pathway is used by plants which are often drought stressed. With these plants, the stomata are closed both day and night. Here as with CAM Cycling there is night time assimilation of respiratory CO2. The benefit here is that metabolism continues during severe drought. These plants usually have a rapid response to rain showers.
Plants using the last three CAM types are usually found in areas where water shortage alternates with water excess. Epiphytes and lithophytes also use these pathways. Often the benefit of continued metabolism (survival) is at the expense of quantum yield (growth).
Plants which can switch photosynthetic pathways between CAM and C3 depend on environmental factors for the switch e.g. plants under water stress will switch to CAM as will plants under saline stress. Plants which are dry then exposed to moisture switch to C3. Note some C4 plants can switch to CAM (no bromeliads use the C4 pathway). Some plants express CAM in their stems and branches. With CAM photorespiration is limited, transpiration is limited and so water efficiency is at least five times greater than for C3 and C4 plants.
CAM Plants and Their Characteristics
| * Of the vascular plant species, some 7% or 15,000-20,000 species, 300 genera, and 40 families are CAM plants (this is considered an underestimate). As stated previously the majority of CAM plants are either epiphytes or succulents, although just about every other growing environment is represented. Most are angiosperms (flowering plants), and CAM species are five times more numerous than C4 species. There are a number of factors which influence the degree of CAM photosynthetic pathway, and these include salinity; pollutants, these decrease the nocturnal CO2 uptake; nutrient availability; increased CO2, which increases the malate; the light level; oxygen; air vapour pressure; temperature; water stress, which influences the enzyme type and volume; nitrogen etc. |
| * CAM plants often show xerophytic characters which include; thickened and reduced leaves, which have a low surface to volume ratio; thickened cuticles; sunken stomata; trichomes; and many CAM plants shed their leaves in the dry season. |
| * Because of the controlled use of CO2 and water, the photosynthetic process is protected from CO2 and water stresses; few other plants can survive such extended neglect ... my kind of plant. |
| * CAM plants can separate the photosynthetic light and dark processes. |
| * Large vacuoles; reduced intercellular air-space; increased cell size. |
| * Because the CAM primary driver is the frugal use of water, CAM plants have meagre photosynthetic rates, and hence suffer a yield (growth) pen- alty. CAM plants need more energy to fix CO2 than C3 or C4 plants. C4 plants have the highest growth rate of all land plants, whereas CAM plants are amongst the slowest growing on earth. C3 plants grow predominantly at night, but CAM plants maximum growth rate is in the middle of the day. |
| * Net CO2 exchange is inhibited by surface wetting. This is a clue on when not to water your CAM broms, since exchange occurs at night. |
| * The more the stress the higher the usage of CO2 recycling, so that the photosynthetic process is little affected by drought. |
| * CAM plants fix CO2 15% more efficiently than C3 plants, but 10% less efficiently than C4 plants. |
| * The CAM pathway involves a temporal concentration of CO2 around the RubisCo enzyme, whereas the C4 pathway involves a spacial concentration of CO2 about RuBisCo. |
Bromeliaceae
69% of the Bromeliaceae are CAM plants or CAM-C3 (meaning depending on the conditions can convert to either). Obviously then, 31% are C3 plants. There are no C4 plants in this family. The table highlights which broms are CAM within the family.
Puya 24% Dyckia and relatives 100% Hectia 100% Tillandsioideae 28% nearly all the atmospherics | |
Note that the Orchidaceae has more CAM species than any other plant family. As a generalization, those bromeliads which are atmospheric Tillandsias, or tank bromeliads with trichomes and stiff leaves are CAM plants. C3 plants have softer leaves and live in shaded and less stressful habitats. However there are many exceptions and the photosynthetic pathway is difficult to identify with morphology alone. The experts have done it by identifying the prevalent enzyme (major carboxylating agent) in the brom. RuBisCo for C3 broms and PEP for CAM broms.
CAM is a means for successful colonization of different habitats, particularly the stressful habitats such as arid, sandy, salty, rocky, and high and low light, together with the habitats of epiphytes and lithophytes. It is probable that CAM is more of a survival mechanism than a biomass increaser. CAM is enhanced by drought.
A couple of interesting points regarding CAM broms, are that water on the leaves appears to prevent the uptake of CO2 because the trichomes become
bloated and flattened and block the stomata. Also the leaves contain a pigment called zeaxanthin that prevents photo-damage (sunburn) to the photosynthesis
apparatus.
Evolution
CAM has evolved convergently many times i.e. the same biological trait is the end result in different or unrelated lineages. In the Bromeliaceae it has evolved at least four times in response to climatic and geologic changes since the late Tertiary ( 2.5 million years).
Within the subfamily Tillandsioideae C3 is plesiomorphic (ancestral) and CAM has developed later in most extreme epiphytes. In the subfamily Bromelioideae
CAM predates epiphytism with subsequent radiation into less xeric habitats and with reversion to C3 in some taxa. Thus we have gained and lost CAM in evolutionary history. The evolutionary trend, terrestrial to epiphytic is closely linked to the elaboration of absorptive epidermal trichomes that are characteristic of the family. CAM broms come in all shapes and sizes, i.e. they are extremely diversified, from soil rooted terrestrials to rosulate tank broms which impound both water and nutrients, to rootless extreme epiphytes which are independent of the substrate.
To sort out a more precise evolution of CAM within the Bromeliaceae one needs a robust phylogeny (evolution) for the family based on molecular (genetic) and morphological characteristics, something which needs more work and is unavailable at present. Consequently many taxonomic relationships remain controversial. Since it is not possible to assign precise chronology to the family’s history it is equally impossible to construct the history for CAM in the Bromeliaceae. However one thing is clear, and that is CAM is a ‘Key Innovation’ associated with the success of broms and their adaptive radiation into more xeric (arid) habitats.
The Bromeliaceae are relatively young but almost completely absent from the fossil record. There is a single report of a Tillandsia type pollen from the upper Eocene (approx. 35 m.y.). Because this is a fairly dubious piece of evidence, scientists have reverted to other means to establish a beginning and develop a history for the Bromeliaceae. Because of the neotropical distribution of broms the conclusion is drawn that the beginning must have come some time after the western Gondwana break-up, and with the separation of South America and Africa sufficient to prevent biological exchange (approx. 85m.y.). There are plant fossils in other families related to the Bromeliaceae (Order Poales), perhaps also the Bromeliaceae emerged at this time in the early Tertiary (65 m.y.). All this is inconclusive and no date of origin or family history for the Bromeliaceae has as yet been established. Thus far it is all surmise.
However some help is gained by the mainly Andean distribution of Puya and the abundance of Tillandsioideae in northern Peru, Equador and Colombia suggesting diversification and radiation into new habitats formed during the Andean mountain building episodes from the Miocene to the Pliocene (23-2.5 m.y.). Certainly the declining concentration of CO2 in the Tertiary would have favoured the emergence of the CAM pathway in broms, as it did for the C4 pathway.
It is appropriate at this stage to mention the remarkable epiphyte Guzmania monostachia. Appropriate because the plant may have evolutionary implications, and remarkable because it has an intermediate photosynthetic pathway between C3 and CAM Idling. There are other species of other genera which may possess this trait but as yet they are undocumented. Guz. monostachia when well watered is a C3 plant and when confronted with drought conditions reverts to the CAM Idling pathway. Suffice it to say there are functional differences along the length of its leaves and resultant divisions of labour which aid this process. CAM Idling is induced by drought stress very quickly (after seven days verses 150 days for an Aechmea species) and since this extremely efficient pathway is seen as a survival mechanism, we have one special plant.
CAM Benefits
| * The shutting of the stomata during the day leads to greater water efficiency. This is particularly useful for seasonal and intermittent water supply. |
| * The CAM pathway keeps the metabolism going in stressful conditions. This is a survival mechanism rather than a biomass or growth producer. |
| * The pathway provides maximum CO2 uptake, minimum photorespiration, and minimum transpiration. |
| * There appear to be four CAM clades (a single ancestor and all its descendants), in the Bromeliaceae, which all have greater species richness and diversity than the C3 clades. |
| * CAM plants are very tough and can survive extreme conditions leading to successful colonization of different habitats. They are very competitive and cling to keeping the metabolic processes alive. |
| * CAM is the first case of a physiological attribute being a ‘Key Innovation’ in plants.i.e. evolution of the CAM photosynthetic pathway and the ensuing colonization of arid and other extreme environments, has promoted taxonomic diversification in the Bromeliaceae. |
References:
In an attempt to explain the CAM photosynthetic pathway in mostly layman's terms (some technical terms are unavoidable), the article comprises information from the following scientific articles and internet pages. Just reinventing the wheel.
Black C. C. and Osmond C.B. 2003. Crassulacean acid metabolism photosynthesis: ‘working the night shift'. Photosynthesis Research 76: 329–341.
Borland A.M. et al 2011. The photosynthetic plasticity of crassulacean acid metabolism: an evolutionary innovation for sustainable productivity in a changing world. New Phytologist 191: 619–633.
Crayn D.M. et al 2004. Multiple origins of crassulacean acid metabolism and the epiphytic habit in the Neotropical family Bromeliaceae. PNAS .101 No.10: 3703–3708.
Cushman J.C. and Borland A.M. 2002. Induction of Crassulacean acid metabolism by water limitation. Plant, Cell and Environment 25, 295–310.
Freschi L. et al 2010. Specific leaf areas of the tank bromeliad Guzmania monostachia perform distinct functions in response to water shortage. Journal of Plant Physiology 167: 526–533.
Haslam R. et al 2003. Physiological responses of the CAM epiphyte Tillandsia usneoides L. (Bromeliaceae) to variations in light and water supply. J. Plant Physiol. 160: 627–634.
Martin C.E. 1994. Physiological Ecology of the Bromeliaceae. Botanical Review, Vol. 60, No.1: 82
Nelson E.A.and Sage R. F. 2008. Functional constraints of CAM leaf anatomy: tight cell packing is associated with increased CAM function across a gradient of CAM expression. Journal of Experimental Botany, Vol. 59, No. 7: 1841–1850.
Nobel P.S. 1991. Achievable productivities of certain CAM plants: basis for high values compared with C3 and C4 plants. New Phytol. 119: 183-205.
Pierce S. et al 2002. The role of CAM in high rainfall cloud forests: an in situ comparison of photo synthetic pathways in Bromeliaceae. Plant, Cell and Environment 25, 1181–1189.
Quezada I. M and Gianoli E. 2011. Crassulacean acid metabolism photosynthesis in Bromeliaceae: an evolutionary key innovation. Biological Journal of the Linnean Society, 104: 480–486.
www.ag.unr.edu/cam/education.aspx
www.en.wikipedia.org/wiki/Crassulacean_acid_metabolism
www.plantsinaction.science.uq.edu.au/edition1/?q=content/2-1-6-crassulacean-acid-metabolism-cam
www.marietta.edu/~biol/biomes/photosynthesis.htm
www.wc.pima.edu/~bfiero/tucsonecology/plants/plants_photosynthesis.htm
Ed: Another comprehensive study of photosynthesis can be found at www.fragrancex.com/fragrance-information/photosynthesis-in-plants-and-flowers.html
